研究紹介
概要




生殖細胞は個体の遺伝情報を伝える細胞であり、体細胞は生殖細胞の乗り物に過ぎません。生殖細胞の中でも増殖する能力をもつものは発生段階の始原生殖細胞と生後の精巣にある精原細胞に限られます。とくに後者の集団中にごく少数存在する精子幹細胞は自己複製能をもち無限に増殖する能力があります。個体は子孫へと遺伝子情報を伝えるためにさまざまな工夫をこらします。神経や免疫系の発達などが起こるのも、個体がより多くの子孫へと効率良く遺伝情報を残すための仕組みだと考えることもできます。生殖細胞の中で唯一自己複製能をもつ精子幹細胞は、生殖細胞の中でも最上位に属し、ある意味では体を構成する細胞の中で最も「利己的な」細胞ではないでしょうか。精子幹細胞が絶え間なく自己複製を繰り返す過程で起こった遺伝子変異は進化や病気の原因となります。これだけの生物学的な重要性をもつにも関わらず、この細胞の実体についてはほとんど情報がありません。
移植精巣
精子幹細胞の研究は長らく形態学的記載にとどまっていましたが、 1994年に米国ペンシルバニア大学のブリンスター博士が精子幹細胞移植法を開発したことで大きく状況が変わりました (Brinster RL, Zimmermann JW, Proc Natl Acad Sci USA 1994;91:11298-11302)。 この方法では薬剤処理により不妊になったホストの精巣の精細管内に、ドナー由来のバラバラにした精巣細胞を注入すると、移植された細胞の中に存在する精子幹細胞がホストの精巣内で精子に分化し、ドナー由来の子孫を作るというものです。この移植法の開発には二つの重要な意味がありました。第一に、これまで移植による幹細胞の機能的アッセイは血液幹細胞でのみ可能でしたが、同じような研究手法と概念で精子幹細胞の解析をすることができるようになったことです。第二に、精子幹細胞は幹細胞の中でも「子孫に遺伝情報を伝達できる」という特殊な能力を持っているため、遺伝子改変動物作成に利用できる可能性が示されました。
私たちのグループは2003年にマウスの精子幹細胞を試験管内で増幅する培養系を確立しました(Kanatsu-Shinohara M.et al., Biol Reprod 2003;69:612-616)。この培養では精子幹細胞の自己複製因子Glial cell line derived neurotrophic factor (GDNF) を含有する低血清培地で精巣細胞を培養すると、精子幹細胞のコロニー形成がみられ、2年以上安定して増殖します。私たちはこの細胞をGermline Stem (GS)細胞と命名しました。GS細胞の樹立により生殖細胞を試験管内で増やすことが初めて可能になりました。
GS細胞は精原細胞マーカーを発現、培養中で増やした後に精巣に戻すと精子へと分化して、産仔を作ることができます。この細胞は安定性が高く、2年間の連続培養後も正常な核型とインプリンティングパターンを保ち、子孫作成能を維持しています(Kanatsu-Shinohara M. et al., Development 2005;132:4155-4163)。ES細胞では数ヶ月にわたる継代とともに染色体(特に8、11番染色体のトリソミー)とDNAメチル化の異常が蓄積し、生殖細胞への分化能を容易に失うとされていますので、生後の幹細胞には幹細胞特有の安全機構が備わっているのではないかと考えています。
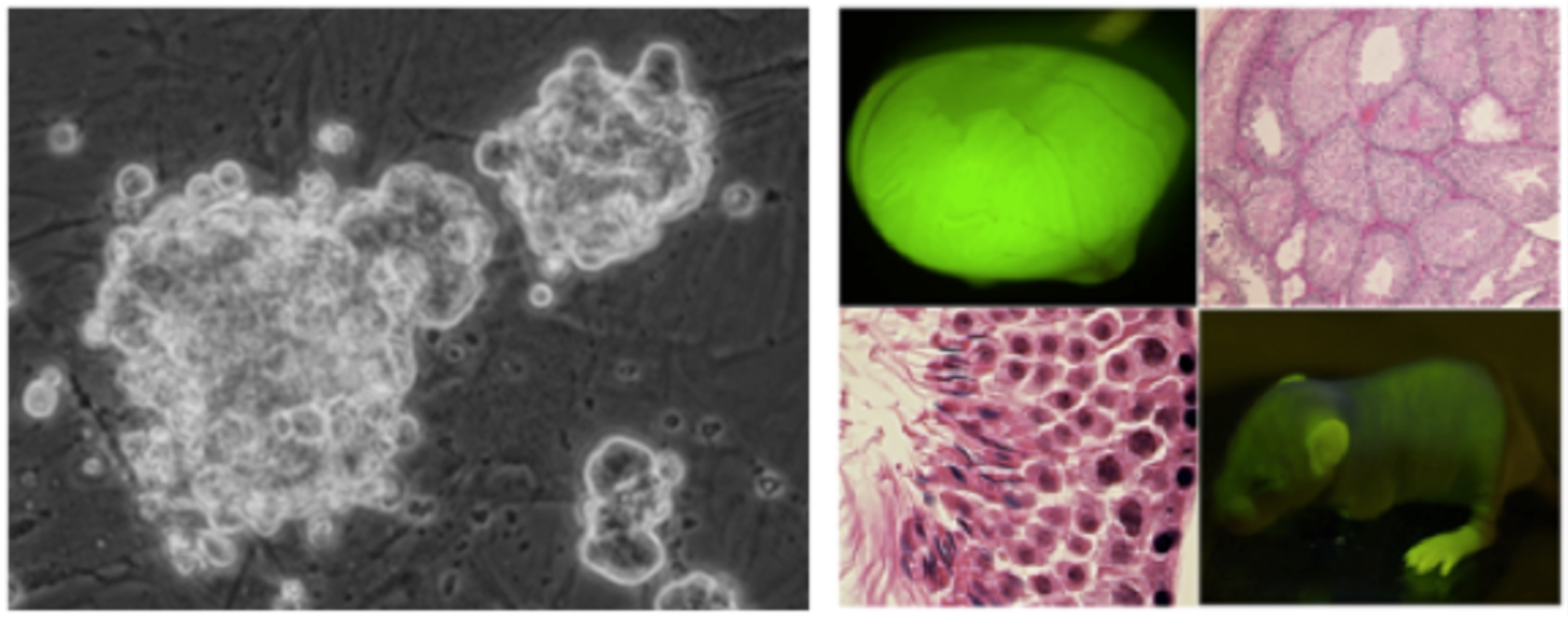
(図の説明)GS細胞: mouse embryonic fibroblast (MEF)フィーダー上にてぶどうの房状のコロニーを作って長期にわたり精原細胞として増殖する(左) 二年間の培養後も精巣内移植により子孫を作る(右)
精巣における精子幹細胞の頻度は非常に低く解析が困難でしたが、GS細胞培養系を用いることで、試験管内操作により大量の幹細胞を得ることが容易になりました。また試験管内で遺伝子操作したGS細胞を精巣に移植して遺伝子改変動物を作ることもできるので、畜産や創薬への技術応用やヒトの不妊治療への応用も期待されています。また、GS細胞は潜在的な多能性を持っており、ES細胞と同等の多能性をもつmultipotent GS (mGS) 細胞にも変化することができます(Kanatsu-Shinohara M.et al., Cell 2004;119:1001-1012.)。 配偶子になり得る培養細胞はこれまでES細胞しかありませんでしたが、GS細胞の樹立により新しい世界が見えてきました。
現在、私たちの教室では①自己複製メカニズムの解明、②精子幹細胞を用いた遺伝子改変動物作成技術の開発(試験管内精子形成)、③子孫への遺伝子伝達パターンの解析、④老化とDNA修復機構の解明、⑤多能性抑制のメカニズムの解明などを行っています。将来的にはこの細胞をES細胞のかわりに個体遺伝子操作の道具として幅広い種でつかえるように開発していこうと考えています。(これまでの仕事のまとめは(Kanatsu-Shinohara M, Shinohara T, Annu Rev Cell Dev Biol 2013;29;163-187)を参照してください)。